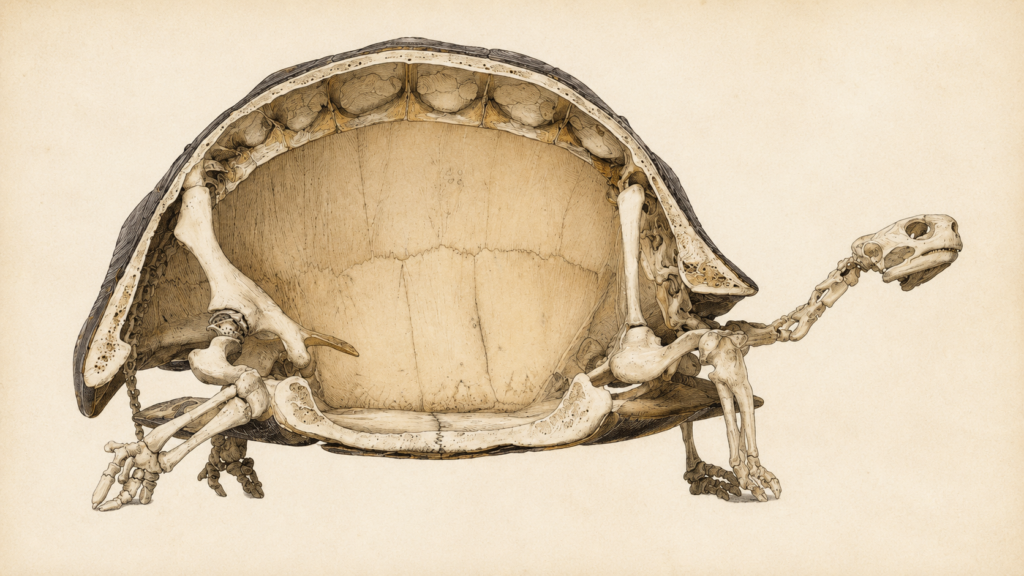
Der Körperbau der Schildkröten ist einzigartig im Tierreich. Kein anderes Wirbeltier besitzt eine vergleichbare Kombination aus innerem Skelett und schützendem Panzer. Diese besondere Anatomie ist das Ergebnis einer langen evolutionären Entwicklung und bildet die Grundlage für die außergewöhnliche Anpassungsfähigkeit dieser Tiere.
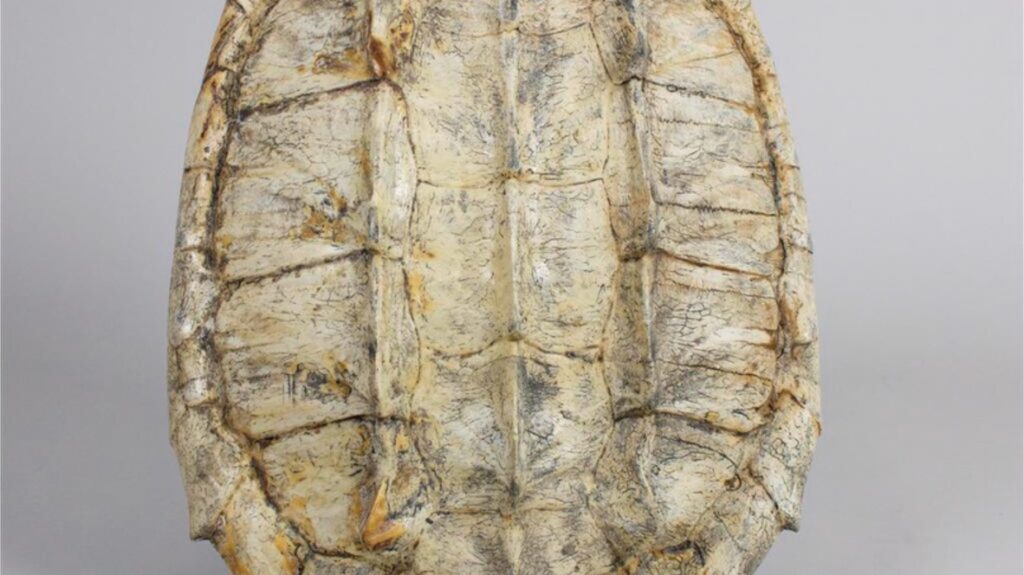
Anatomie des Schildkrötenpanzers: Einzigartig unter Wirbeltieren
Der Panzer der Schildkröte ist keine bloße „Schachtel“, sondern ein hochentwickelter, evolutionär einzigartiger Teil des Skeletts. Seine Anatomie unterscheidet diese Reptilien fundamental von allen anderen Wirbeltieren.
Der anatomische Bauplan
- Zweiteiliger Schutz: Der Panzer besteht aus dem gewölbten Rückenpanzer (Carapax) und dem flachen Bauchpanzer (Plastron).
- Feste Knochenbrücke: Beide Panzerteile sind an den Seiten starr miteinander verschmolzen.
- Teil des Innenskeletts: Der Panzer ist kein totes Außenskelett. Er besteht aus lebendigem Knochengewebe.
- Verschmolzene Knochen: Wirbelsäule und Rippen sind direkt in die Knochenplatten des Panzers hineingewachsen.
- Innere Schultern: Der Schultergürtel liegt paradoxerweise innerhalb des Rippenkorbs. Dies ist einmalig im Tierreich.
Die äußere Schicht
- Hornschilde: Die meisten Arten tragen eine Oberschicht aus Keratinplatten (Horn), ähnlich unseren Fingernägeln.
- Lederschicht: Manche Arten (wie Weichschildkröten) besitzen stattdessen eine dicke, lederartige Haut.
- Form folgt Funktion: Flache Panzer optimieren die Stromlinienform im Wasser. Hochgewölbte Panzer schützen Landschildkröten vor Raubtiergebissen.
Anpassungen an Atmung und Bewegung
Der starre Panzer bringt funktionelle Herausforderungen mit sich. Da sich der Brustkorb nicht ausdehnen kann, erfolgt die Atmung über spezielle Muskelbewegungen, die mit den Extremitäten gekoppelt sind.
Auch die Fortbewegung ist eng mit dem Körperbau verknüpft. An Land bewegen sich Schildkröten schreitend mit kräftigen Beinen, während aquatische Arten rudernde Bewegungen nutzen. Meeresschildkröten haben ihre Vorderextremitäten zu flossenartigen Strukturen umgebildet, die ihnen effizientes Schwimmen über große Distanzen ermöglichen.
Wie Schildkröten ihre Umwelt wahrnehmen
Sehvermögen und UV-Wahrnehmung
Für Schildkröten ist Farbwahrnehmung einschließlich UV-Anteilen gut belegt. Bei 𝘗𝘴𝘦𝘶𝘥𝘦𝘮𝘺𝘴 𝘴𝘤𝘳𝘪𝘱𝘵𝘢 𝘦𝘭𝘦𝘨𝘢𝘯𝘴 wurde ein vierter, UV-sensitiver Zapfentyp nachgewiesen; die maximale UV-Empfindlichkeit lag bei etwa 370–380 nm. Weitere Arbeiten zeigen UV-Eingänge in der Retina und UV-Farbgegensätze in retinalen Nervenzellen.

Geruchssinn und chemische Orientierung
Für Meeresschildkröten ist belegt, dass die Nasenhöhle an Wasserfluss und Wasserolfaktion angepasst ist. Neuere anatomische Arbeiten an 𝘊𝘩𝘦𝘭𝘰𝘯𝘪𝘢 𝘮𝘺𝘥𝘢𝘴 zeigen, dass sowohl flüchtige als auch wasserlösliche Duftstoffe über spezialisierte Strukturen der Nasenhöhle wahrgenommen werden können. Bei 𝘊𝘢𝘳𝘦𝘵𝘵𝘢 𝘤𝘢𝘳𝘦𝘵𝘵𝘢 wurde zudem die Wahrnehmung von Luftgerüchen experimentell nachgewiesen.
Hören und tieffrequente Reize
Das Gehör von Schildkröten ist besonders für niedrigere Frequenzen relevant. Bei juvenilen Grünen Meeresschildkröten wurden Reaktionen auf Unterwasserschall zwischen 50 und 1.600 Hz gemessen, mit höchster Empfindlichkeit zwischen 200 und 400 Hz. Die Autoren betonen außerdem, dass Schalldetektion sowohl unter Wasser als auch in der Luft möglich ist, die biologische Bedeutung aber noch nicht vollständig verstanden ist.
Vibrationen
Für Schildkröten sind akustische und vibrationale Reize eng miteinander verknüpft. Arbeiten zur Meeresschildkrötenakustik verweisen darauf, dass sowohl Luft- bzw. Wasserschall als auch Vibrationen über das Hörsystem verarbeitet werden können; besonders niedrigfrequente Reize sind dabei relevant.
Räumliches Gedächtnis und Lernfähigkeit
Räumliches Lernen ist bei Schildkröten gut untersucht. In Labyrinthversuchen mit 𝘗𝘴𝘦𝘶𝘥𝘦𝘮𝘺𝘴 𝘴𝘤𝘳𝘪𝘱𝘵𝘢 zeigten die Tiere sowohl ortsbezogenes Lernen als auch cue-basiertes Lernen. Bei der Köhler-Schildkröte 𝘊𝘩𝘦𝘭𝘰𝘯𝘰𝘪𝘥𝘪𝘴 𝘤𝘢𝘳𝘣𝘰𝘯𝘢𝘳𝘪𝘶𝘴, wurden räumliches Lernen und das Erinnern an bereits besuchte bzw. belohnte Orte nachgewiesen.
Kommunikation mit Lauten, Gerüchen und Körpersprache
Lange galten Schildkröten als weitgehend stumme Tiere. Moderne verhaltensbiologische und bioakustische Studien zeigen jedoch, dass viele Arten Laute erzeugen und diese in bestimmten sozialen Zusammenhängen nutzen können. Besonders gut belegt sind Unterwasserlaute bei verschiedenen Wasserschildkröten sowie Balzlaute bei Landschildkröten. Bei einigen Arten wurden sogar Lautäußerungen aus dem Ei oder aus dem Nest dokumentiert, wobei ihre genaue Funktion je nach Art noch nicht vollständig geklärt ist.
Akustische Kommunikation als uraltes Erbe
Die Lautäußerungen von Schildkröten sind nicht nur verhaltensbiologisch interessant, sondern auch evolutionsgeschichtlich bedeutsam. Lange wurden viele Reptilien, Amphibien und andere ursprüngliche Wirbeltiergruppen als weitgehend stumm betrachtet. Neue bioakustische Untersuchungen zeigen jedoch, dass gerade solche wenig beachteten Tiergruppen ein deutlich breiteres Lautrepertoire besitzen als früher angenommen.
Eine große vergleichende Studie kam zu dem Ergebnis, dass akustische Kommunikation bei Wirbeltieren möglicherweise nicht mehrfach unabhängig entstanden ist, sondern auf einen sehr alten gemeinsamen Ursprung zurückgehen könnte. Die Fähigkeit, Laute zur Verständigung einzusetzen, wäre demnach bereits beim letzten gemeinsamen Vorfahren der Choanata vorhanden gewesen, also bei einer Linie, zu der Lungenfische und Landwirbeltiere gehören. Dieser Vorfahr lebte vor rund 407 Millionen Jahren. Schildkröten spielen in dieser Rekonstruktion eine wichtige Rolle, weil sie eine bisher unterschätzte Datenlücke schließen: Ihre Laute zeigen, dass auch vermeintlich „stumme“ Tiergruppen wichtige Hinweise auf die frühe Evolution der Kommunikation liefern können. Die Befunde deuten darauf hin, dass akustische Kommunikation bei Wirbeltieren wesentlich älter sein könnte als lange angenommen.
Kommunikation bei Schildkröten ist jedoch nicht auf Laute beschränkt. Geruchssignale, Körperhaltung, Kopfnicken, Umrunden, Rammen, Beißen und taktile Reize bilden zusammen ein multimodales Kommunikationssystem. Vor allem bei Balz, Partnererkennung, Rivalität und sozialer Orientierung greifen akustische, chemische und visuelle Signale ineinander.
Vielfalt der Anpassungen
Die Kombination aus Panzer, Sinnesleistungen und spezialisierten Bewegungsformen ermöglicht es Schildkröten, unterschiedlichste Lebensräume zu besiedeln. Von wüstenbewohnenden Landschildkröten bis hin zu hochspezialisierten Meeresschildkröten zeigt sich eine enorme Bandbreite an Anpassungen.
Diese Vielfalt macht den Körperbau der Schildkröten zu einem der faszinierendsten Beispiele evolutionärer Innovation.
Weiterführende Themen
Grundlagen im Artikel „Schildkröten – Evolution und Vielfalt“
Gefährdung im Artikel „Taxonomie und Bedrohung“
Ernährung im Artikel „Ernährung und Verhalten“
Artenschutz-Instrumente im Artikel „Die Wächter der Vielfalt„
Bemerkenswertes im Artikel „Besondere Fakten„
Thermoregulation der Lederschildkröten in „Gigantothermie„
Anatomisches in „Körperbau“
„Entstehung“ der mediterranen Landschildkröten
„Refugien„ während der Eiszeit
Quellen
AMMERMÜLLER, J., ITZHAKI, A., WEILER, R. & PERLMAN, I. (1998): UV-sensitive input to horizontal cells in the turtle retina. European Journal of Neuroscience 10(5): 1544–1552.
ARNOLD, K. & NEUMEYER, C. (1987): Wavelength discrimination in the turtle 𝘗𝘴𝘦𝘶𝘥𝘦𝘮𝘺𝘴 𝘴𝘤𝘳𝘪𝘱𝘵𝘢 𝘦𝘭𝘦𝘨𝘢𝘯𝘴. Vision Research 27(9): 1501–1511.
BARTOL, S. M., MUSICK, J. A. & LENHARDT, M. L. (1999): Auditory evoked potentials of the loggerhead sea turtle 𝘊𝘢𝘳𝘦𝘵𝘵𝘢 𝘤𝘢𝘳𝘦𝘵𝘵𝘢. Copeia 1999(3): 836–840.
ENDRES, C. S., PUTMAN, N. F. & LOHMANN, K. J. (2009): Perception of airborne odors by loggerhead sea turtles. Journal of Experimental Biology 212(23): 3823–3827.
FERRARA, C. R., VOGT, R. C. & SOUSA-LIMA, R. S. (2013): Turtle vocalizations as the first evidence of posthatching parental care in chelonians. Journal of Comparative Psychology 127(1): 24–32.
GALEOTTI, P., SACCHI, R., FASOLA, M. & BALLASINA, D. (2005): Do mounting vocalisations in tortoises have a communication function? A comparative analysis. Herpetological Journal 15(1): 61–71.
GALEOTTI, P., SACCHI, R., PELLITTERI-ROSA, D. & FASOLA, M. (2007): Olfactory discrimination of species, sex, and sexual maturity by the Hermann’s tortoise 𝘛𝘦𝘴𝘵𝘶𝘥𝘰 𝘩𝘦𝘳𝘮𝘢𝘯𝘯𝘪. Copeia 2007(4): 980–985.
GILES, J. C., DAVIS, J. A., MCCAULEY, R. D. & KUCHLING, G. (2009): Voice of the turtle: The underwater acoustic repertoire of the long-necked freshwater turtle 𝘊𝘩𝘦𝘭𝘰𝘥𝘪𝘯𝘢 𝘰𝘣𝘭𝘰𝘯𝘨𝘢. The Journal of the Acoustical Society of America 126(1): 434–443.
IBÁÑEZ, A., FRITZ, U., AUER, M., MARTÍNEZ-SILVESTRE, A. & PABIJAN, M. (2021): Evolutionary history of mental glands in turtles reveals a single origin in an aquatic ancestor and recurrent losses independent of macrohabitat. Scientific Reports 11: 10396.
JORGEWICH-COHEN, G., TOWNSEND, S. W., PADOVESE, L. R., KLEIN, N., PRASCHAG, P., FERRARA, C. R., ETTMAR, S., MENEZES, S. et al. (2022): Common evolutionary origin of acoustic communication in choanate vertebrates. Nature Communications 13: 6089.
JORGEWICH-COHEN, G., WHEATLEY, M., GASPAR, L. P., PRASCHAG, P., LUBBERINK, N. S., MING, K., RODRIGUEZ, N. A. & FERRARA, C. R. (2024): Prehatch calls and coordinated birth in turtles. Ecology and Evolution 14(10): e70410.
KELLEY, M. D., KA, C., FINGER, J. W. & MENDONÇA, M. T. (2021): Behavioural discrimination of male mental gland secretions of the gopher tortoise 𝘎𝘰𝘱𝘩𝘦𝘳𝘶𝘴 𝘱𝘰𝘭𝘺𝘱𝘩𝘦𝘮𝘶𝘴 by both sexes. Behavioural Processes 183: 104314.
KELLEY, M. D., FINGER, J. W. & MENDONÇA, M. T. (2022): Male gopher tortoise 𝘎𝘰𝘱𝘩𝘦𝘳𝘶𝘴 𝘱𝘰𝘭𝘺𝘱𝘩𝘦𝘮𝘶𝘴 concentration-dependent social responses to diluted mental gland pheromones. Behavioural Processes 201: 104729.
KONDOH, D., KITAYAMA, C. & KAWAI, Y. K. (2021): The nasal cavity in sea turtles: adaptation to olfaction and seawater flow. Cell and Tissue Research 383(1): 347–352.
KONDOH, D., KANEOYA, Y., TONOMORI, W. & KITAYAMA, C. (2023): Histological features and Gαolf expression patterns in the nasal cavity of sea turtles. Journal of Anatomy 243(3): 486–503.
LÓPEZ, J. C., RODRÍGUEZ, F., GÓMEZ, Y., VARGAS, J. P., BROGLIO, C. & SALAS, C. (2000): Place and cue learning in turtles. Animal Learning & Behavior 28: 360–372.
MASON, R. T. & PARKER, M. R. (2010): Social behavior and pheromonal communication in reptiles. Journal of Comparative Physiology A 196: 729–749.
MCGLASHAN, J. K., SPENCER, R.-J. & OLD, J. M. (2012): Embryonic communication in the nest: metabolic responses of reptilian embryos to developmental rates of siblings. Proceedings of the Royal Society B 279: 1709–1715.
PINIAK, W. E. D., MANN, D. A., HARMS, C. A., JONES, T. T. & ECKERT, S. A. (2016): Hearing in the juvenile green sea turtle 𝘊𝘩𝘦𝘭𝘰𝘯𝘪𝘢 𝘮𝘺𝘥𝘢𝘴: A comparison of underwater and aerial hearing using auditory evoked potentials. PLOS ONE 11(10): e0159711.
SACCHI, R., GALEOTTI, P., FASOLA, M. & BALLASINA, D. (2003): Vocalizations and courtship intensity correlate with mounting success in marginated tortoises 𝘛𝘦𝘴𝘵𝘶𝘥𝘰 𝘮𝘢𝘳𝘨𝘪𝘯𝘢𝘵𝘢. Behavioral Ecology and Sociobiology 55: 95–102.
VENTURA, D. F., ZANA, Y., DE SOUZA, J. M. & DEVOE, R. D. (2001): Ultraviolet colour opponency in the turtle retina. Journal of Experimental Biology 204(14): 2527–2534.
WILKINSON, A., CHAN, H.-M. & HALL, G. (2007): Spatial learning and memory in the tortoise 𝘎𝘦𝘰𝘤𝘩𝘦𝘭𝘰𝘯𝘦 𝘤𝘢𝘳𝘣𝘰𝘯𝘢𝘳𝘪𝘢. Journal of Comparative Psychology 121(4): 412–418.
WILKINSON, A., COWARD, S. & HALL, G. (2009): Visual and response-based navigation in the tortoise 𝘎𝘦𝘰𝘤𝘩𝘦𝘭𝘰𝘯𝘦 𝘤𝘢𝘳𝘣𝘰𝘯𝘢𝘳𝘪𝘢. Animal Cognition 12: 779–787.
The Anatomy of Turtles and Tortoises – Shell and Senses
The anatomy of turtles and tortoises is unique in the animal kingdom. No other vertebrate has a comparable combination of internal skeleton and protective shell. This distinctive anatomy is the result of a long evolutionary history and forms the basis of these animals’ remarkable adaptability.
The bony shell forms a single unit with the turtle’s spine.
The shell – a unique structural design
The most striking feature of all turtles and tortoises is their shell. It consists of two main parts: the upper shell, or carapace, and the lower shell, or plastron. These two parts are firmly connected and enclose a large part of the body.
Unlike an external skeleton, the shell is part of the internal skeleton. It evolved from the ribs, vertebrae and bony plates. This fusion makes the chelonian shell a unique structure among vertebrates.
One particularly distinctive anatomical feature is the position of the shoulder girdle, which lies inside the ribcage. This unusual body plan fundamentally distinguishes turtles and tortoises from all other vertebrates.
Depending on the species, the shell is covered with keratinous scutes or with leathery skin. Its shape, structure and stability vary considerably and reflect the animal’s particular way of life.
Adaptations for breathing and movement
The rigid shell creates functional challenges. Because the ribcage cannot expand, breathing is achieved through specialised muscle movements that are linked to the limbs.
Locomotion is also closely connected with body structure. On land, tortoises move with a walking gait using strong legs, while aquatic species use paddling movements. Sea turtles have modified their forelimbs into flipper-like structures, enabling them to swim efficiently over long distances.
Sensory abilities – underestimated skills
Turtles and tortoises have well-developed sensory organs. Their eyesight is particularly well developed and allows them to perceive a broad colour spectrum, including ultraviolet wavelengths.
The sense of smell plays a central role in finding food and orientation. In many aquatic turtles, scent particles are taken up through characteristic movements of the mouth and throat.
Their hearing is specialised for low-frequency vibrations. Turtles and tortoises mainly perceive vibrations from their surroundings, while high-pitched sounds are of lesser importance.
In addition, many species have a good spatial memory and can remember food sources and escape routes over long periods of time.
Communication and behaviour
For a long time, turtles and tortoises were considered silent animals. Today, we know that they can indeed produce sounds, for example during mating or in stressful situations.
Smells, body posture and movements also play an important role in communication. It is particularly remarkable that embryos can already exchange acoustic signals inside the egg, probably to help synchronise hatching.
A diversity of adaptations
The combination of shell, sensory abilities and specialised forms of movement allows turtles and tortoises to occupy a wide range of habitats. From desert-dwelling tortoises to highly specialised sea turtles, they show an enormous diversity of adaptations.
This diversity makes the anatomy of turtles and tortoises one of the most fascinating examples of evolutionary innovation.
Sources
AMMERMÜLLER, J., ITZHAKI, A., WEILER, R. & PERLMAN, I. (1998): UV-sensitive input to horizontal cells in the turtle retina. European Journal of Neuroscience 10(5): 1544–1552.
ARNOLD, K. & NEUMEYER, C. (1987): Wavelength discrimination in the turtle 𝘗𝘴𝘦𝘶𝘥𝘦𝘮𝘺𝘴 𝘴𝘤𝘳𝘪𝘱𝘵𝘢 𝘦𝘭𝘦𝘨𝘢𝘯𝘴. Vision Research 27(9): 1501–1511.
BARTOL, S. M., MUSICK, J. A. & LENHARDT, M. L. (1999): Auditory evoked potentials of the loggerhead sea turtle 𝘊𝘢𝘳𝘦𝘵𝘵𝘢 𝘤𝘢𝘳𝘦𝘵𝘵𝘢. Copeia 1999(3): 836–840.
ENDRES, C. S., PUTMAN, N. F. & LOHMANN, K. J. (2009): Perception of airborne odors by loggerhead sea turtles. Journal of Experimental Biology 212(23): 3823–3827.
FERRARA, C. R., VOGT, R. C. & SOUSA-LIMA, R. S. (2013): Turtle vocalizations as the first evidence of posthatching parental care in chelonians. Journal of Comparative Psychology 127(1): 24–32.
GALEOTTI, P., SACCHI, R., FASOLA, M. & BALLASINA, D. (2005): Do mounting vocalisations in tortoises have a communication function? A comparative analysis. Herpetological Journal 15(1): 61–71.
GALEOTTI, P., SACCHI, R., PELLITTERI-ROSA, D. & FASOLA, M. (2007): Olfactory discrimination of species, sex, and sexual maturity by the Hermann’s tortoise 𝘛𝘦𝘴𝘵𝘶𝘥𝘰 𝘩𝘦𝘳𝘮𝘢𝘯𝘯𝘪. Copeia 2007(4): 980–985.
GILES, J. C., DAVIS, J. A., MCCAULEY, R. D. & KUCHLING, G. (2009): Voice of the turtle: The underwater acoustic repertoire of the long-necked freshwater turtle 𝘊𝘩𝘦𝘭𝘰𝘥𝘪𝘯𝘢 𝘰𝘣𝘭𝘰𝘯𝘨𝘢. The Journal of the Acoustical Society of America 126(1): 434–443.
IBÁÑEZ, A., FRITZ, U., AUER, M., MARTÍNEZ-SILVESTRE, A. & PABIJAN, M. (2021): Evolutionary history of mental glands in turtles reveals a single origin in an aquatic ancestor and recurrent losses independent of macrohabitat. Scientific Reports 11: 10396.
JORGEWICH-COHEN, G., TOWNSEND, S. W., PADOVESE, L. R., KLEIN, N., PRASCHAG, P., FERRARA, C. R., ETTMAR, S., MENEZES, S. et al. (2022): Common evolutionary origin of acoustic communication in choanate vertebrates. Nature Communications 13: 6089.
JORGEWICH-COHEN, G., WHEATLEY, M., GASPAR, L. P., PRASCHAG, P., LUBBERINK, N. S., MING, K., RODRIGUEZ, N. A. & FERRARA, C. R. (2024): Prehatch calls and coordinated birth in turtles. Ecology and Evolution 14(10): e70410.
KELLEY, M. D., KA, C., FINGER, J. W. & MENDONÇA, M. T. (2021): Behavioural discrimination of male mental gland secretions of the gopher tortoise 𝘎𝘰𝘱𝘩𝘦𝘳𝘶𝘴 𝘱𝘰𝘭𝘺𝘱𝘩𝘦𝘮𝘶𝘴 by both sexes. Behavioural Processes 183: 104314.
KELLEY, M. D., FINGER, J. W. & MENDONÇA, M. T. (2022): Male gopher tortoise 𝘎𝘰𝘱𝘩𝘦𝘳𝘶𝘴 𝘱𝘰𝘭𝘺𝘱𝘩𝘦𝘮𝘶𝘴 concentration-dependent social responses to diluted mental gland pheromones. Behavioural Processes 201: 104729.
KONDOH, D., KITAYAMA, C. & KAWAI, Y. K. (2021): The nasal cavity in sea turtles: adaptation to olfaction and seawater flow. Cell and Tissue Research 383(1): 347–352.
KONDOH, D., KANEOYA, Y., TONOMORI, W. & KITAYAMA, C. (2023): Histological features and Gαolf expression patterns in the nasal cavity of sea turtles. Journal of Anatomy 243(3): 486–503.
LÓPEZ, J. C., RODRÍGUEZ, F., GÓMEZ, Y., VARGAS, J. P., BROGLIO, C. & SALAS, C. (2000): Place and cue learning in turtles. Animal Learning & Behavior 28: 360–372.
MASON, R. T. & PARKER, M. R. (2010): Social behavior and pheromonal communication in reptiles. Journal of Comparative Physiology A 196: 729–749.
MCGLASHAN, J. K., SPENCER, R.-J. & OLD, J. M. (2012): Embryonic communication in the nest: metabolic responses of reptilian embryos to developmental rates of siblings. Proceedings of the Royal Society B 279: 1709–1715.
PINIAK, W. E. D., MANN, D. A., HARMS, C. A., JONES, T. T. & ECKERT, S. A. (2016): Hearing in the juvenile green sea turtle 𝘊𝘩𝘦𝘭𝘰𝘯𝘪𝘢 𝘮𝘺𝘥𝘢𝘴: A comparison of underwater and aerial hearing using auditory evoked potentials. PLOS ONE 11(10): e0159711.
SACCHI, R., GALEOTTI, P., FASOLA, M. & BALLASINA, D. (2003): Vocalizations and courtship intensity correlate with mounting success in marginated tortoises 𝘛𝘦𝘴𝘵𝘶𝘥𝘰 𝘮𝘢𝘳𝘨𝘪𝘯𝘢𝘵𝘢. Behavioral Ecology and Sociobiology 55: 95–102.
VENTURA, D. F., ZANA, Y., DE SOUZA, J. M. & DEVOE, R. D. (2001): Ultraviolet colour opponency in the turtle retina. Journal of Experimental Biology 204(14): 2527–2534.
WILKINSON, A., CHAN, H.-M. & HALL, G. (2007): Spatial learning and memory in the tortoise 𝘎𝘦𝘰𝘤𝘩𝘦𝘭𝘰𝘯𝘦 𝘤𝘢𝘳𝘣𝘰𝘯𝘢𝘳𝘪𝘢. Journal of Comparative Psychology 121(4): 412–418.
WILKINSON, A., COWARD, S. & HALL, G. (2009): Visual and response-based navigation in the tortoise 𝘎𝘦𝘰𝘤𝘩𝘦𝘭𝘰𝘯𝘦 𝘤𝘢𝘳𝘣𝘰𝘯𝘢𝘳𝘪𝘢. Animal Cognition 12: 779–787.